
Birds and Bioenergy within the Americas: A Cross-National, Social–Ecological Study of Ecosystem Service Tradeoffs


 ,
,  , , , , , ,
, , , , , ,
Abstract
:1. Introduction
1.1. Background to Study Area
1.1.1. Feedstocks and Human Communities
1.1.2. Feedstocks and Bird Communities
2. Materials and Methods
2.1. Survey Methods
2.2. Ecological Methods
2.3. Site Description
2.4. Point Counts
2.5. Statistical Analyses
3. Results
3.1. Community Survey Results
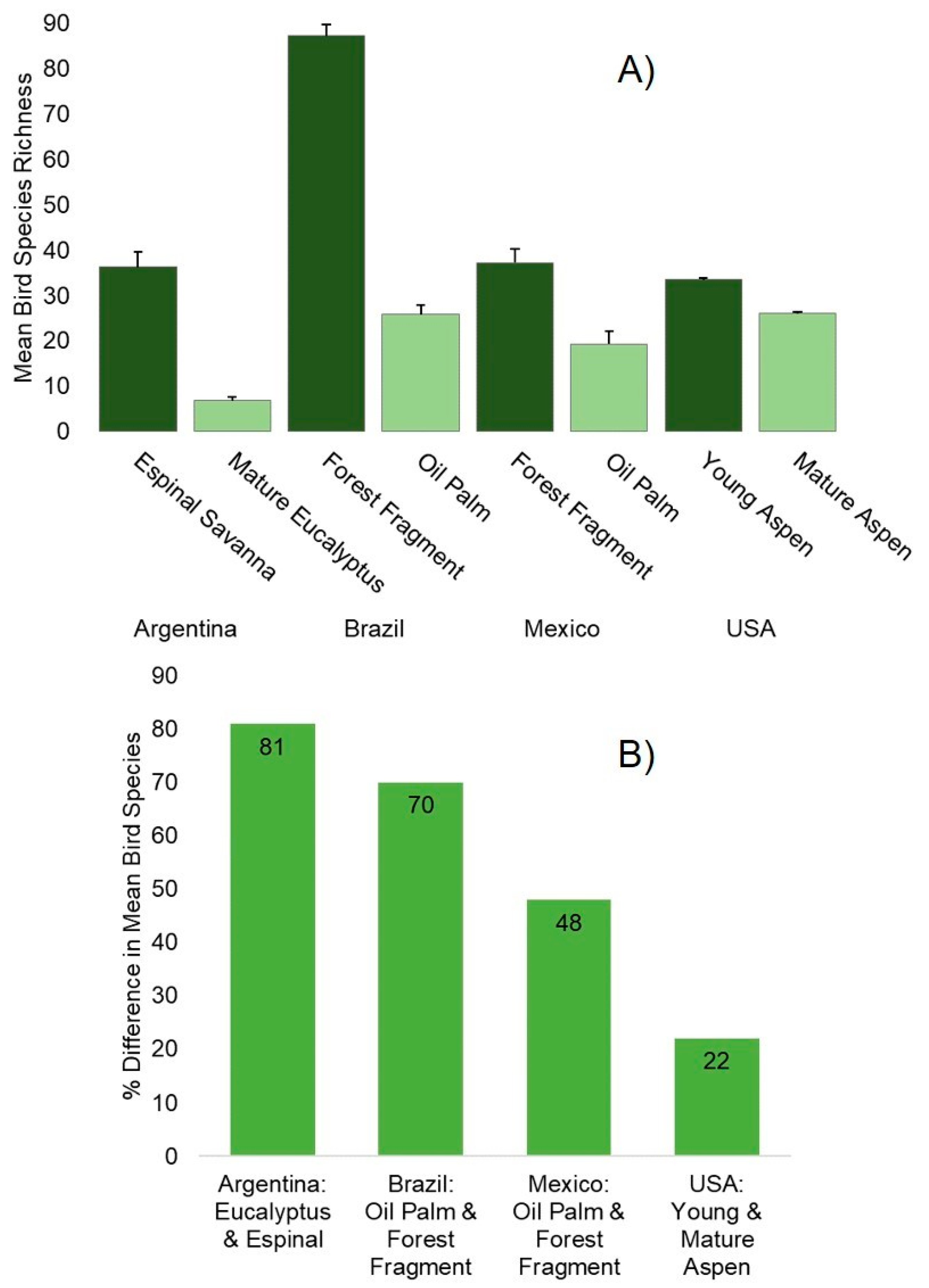
3.2. Ecological Results
3.3. Integrated Social–Ecological Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Owusu, P.A.; Asumadu-Sarkodie, S. A review of renewable energy sources, sustainability issues and climate change mitigation. Cogent Eng. 2016, 3, 1167990. [Google Scholar] [CrossRef]
- Santangeli, A.; Toivonen, T.; Pouzols, F.M.; Pogson, M.; Hastings, A.; Smith, P.; Moilanen, A. Global change synergies and trade-offs between renewable energy and biodiversity. GCB Bioenergy 2016, 8, 941–951. [Google Scholar] [CrossRef] [Green Version]
- Scarlat, N.; Dallemand, J.F.; Monforti-Ferrario, F.; Nita, V. The role of biomass and bioenergy in a future bioeconomy: Policies and facts. Environ. Dev. 2015, 15, 3–34. [Google Scholar] [CrossRef]
- Botelho, A.; Ferreira, P.; Lima, F.; Pinto, L.M.C.; Sousa, S. Assessment of the environmental impacts associated with hydropower. Renew. Sustain. Energy Rev. 2017, 70, 896–904. [Google Scholar] [CrossRef]
- May, R.; Masden, E.A.; Bennet, F.; Perron, M. Considerations for upscaling individual effects of wind energy development towards population-level impacts on wildlife. J. Environ. Manag. 2019, 230, 84–93. [Google Scholar] [CrossRef]
- Felipe-Lucia, M.R.; Soliveres, S.; Penone, C.; Manning, P.; van der Plas, F.; Boch, S.; Prati, D.; Ammer, C.; Schall, P.; Gossner, M.M.; et al. Multiple forest attributes underpin the supply of multiple ecosystem services. Nat. Commun. 2018, 9, 4839. [Google Scholar] [CrossRef] [Green Version]
- Martín-López, B.; Gómez-Baggethun, E.; García-Llorente, M.; Montes, C. Trade-offs across value-domains in ecosystem services assessment. Ecol. Indic. 2014, 37, 220–228. [Google Scholar] [CrossRef]
- Mora, F.; Balvanera, P.; García-Frapolli, E.; Castillo, A.; Trilleras, J.M.; Cohen-Salgado, D.; Salmerón, O. Trade-offs between ecosystem services and alternative pathways toward sustainability in a tropical dry forest region. Ecol. Soc. 2016, 21, 45. [Google Scholar] [CrossRef]
- Mwanukuzi, P.K. Impact of eucalyptus and pine growing on rural livelihood: The lesson from Bukoba area, north western Tanzania. Afr. J. Ecol. 2009, 47, 105–109. [Google Scholar] [CrossRef]
- Phifer, C.C.; Knowlton, J.L.; Webster, C.R.; Flaspohler, D.J.; Licata, J.A. Bird community responses to afforested eucalyptus plantations in the Argentine pampas. Biodivers. Conserv. 2017, 26, 3073–3101. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Runge, C.A.; Gentry, R.R.; Gaines, S.D.; Halpern, B.S. Comparative terrestrial feed and land use of an aquaculture-dominant world. Proc. Natl. Acad. Sci. USA 2018, 115, 5295–5300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krausmann, F.; Erb, K.H.; Gingrich, S.; Haberl, H.; Bondeau, A.; Gaube, V.; Lauk, C.; Plutzar, C.; Searchinger, T.D. Global human appropriation of net primary production doubled in the 20th century. Proc. Natl. Acad. Sci. USA 2013, 110, 10324–10329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of Earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, C. Black is the new green. Conserv. Mag. 2010. Available online: https://www.conservationmagazine.org/2010/08/black-is-the-new-green/ (accessed on 12 December 2018).
- Nájera, A.; Simonetti, J.A. Enhancing Avifauna in Commercial Plantations. Conserv. Biol. 2010, 24, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Perfecto, I.; Vandermeer, J. The agroecological matrix as alternative to the land-sparing/agriculture intensification model. Proc. Natl. Acad. Sci. USA 2010, 107, 5786–5791. [Google Scholar] [CrossRef] [Green Version]
- Pascual, U.; Perrings, C. Developing incentives and economic mechanisms for in situ biodiversity conservation in agricultural landscapes. Agric. Ecosyst. Environ. 2007, 121, 256–268. [Google Scholar] [CrossRef]
- Tomich, T.P.; Brodt, S.; Ferris, H.; Galt, R.; Horwath, W.R.; Kebreab, E.; Leveau, J.H.J.; Liptzin, D.; Lubell, M.; Merel, P.; et al. Agroecology: A Review from a Global-Change Perspective. Annu. Rev. Environ. Resour. 2011, 36, 193–222. [Google Scholar] [CrossRef] [Green Version]
- Mace, G.M.; Norris, K.; Fitter, A.H. Biodiversity and ecosystem services: A multilayered relationship. Trends Ecol. Evol. 2012, 27, 19–26. [Google Scholar] [CrossRef]
- Cox, D.T.C.; Gaston, K.J. Urban Bird Feeding: Connecting People with Nature. PLoS ONE 2016, 11, e0158717. [Google Scholar] [CrossRef] [PubMed]
- Şekercioğlu, C.H. Increasing awareness of avian ecological function. Trends Ecol. Evol. 2006, 21, 464–471. [Google Scholar] [CrossRef]
- Whelan, C.J.; Şekercioğlu, Ç.H.; Wenny, D.G. Why birds matter: From economic ornithology to ecosystem services. J. Ornithol. 2015, 156, 227–238. [Google Scholar] [CrossRef]
- Whelan, C.J.; Wenny, D.G.; Marquis, R.J. Ecosystem Services Provided by Birds. Ann. N. Y. Acad. Sci. 2008, 1134, 25–60. [Google Scholar] [CrossRef] [PubMed]
- Michel, N.L.; Whelan, C.J.; Verutes, G.M. Ecosystem services provided by Neotropical birds. Condor 2020, 122, duaa022. [Google Scholar] [CrossRef]
- Belaire, J.A.; Westphal, L.M.; Whelan, C.J.; Minor, E.S. Urban residents’ perceptions of birds in the neighborhood: Biodiversity, cultural ecosystem services, and disservices. Condor 2015, 117, 192–202. [Google Scholar] [CrossRef]
- Chan, K.M.A.; Guerry, A.D.; Balvanera, P.; Klain, S.; Satterfield, T.; Basurto, X.; Bostrom, A.; Chuenpagdee, R.; Gould, R.; Halpern, B.S.; et al. Where are Cultural and Social in Ecosystem Services? A Framework for Constructive Engagement. Bioscience 2012, 62, 744–756. [Google Scholar] [CrossRef]
- Dallimer, M.; Irvine, K.N.; Skinner, A.M.J.; Davies, Z.G.; Rouquette, J.R.; Maltby, L.L.; Warren, P.H.; Armsworth, P.R.; Gaston, K.J. Biodiversity and the Feel-Good Factor: Understanding Associations between Self-Reported Human Well-being and Species Richness. Bioscience 2012, 62, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Shwartz, A.; Turbé, A.; Simon, L.; Julliard, R. Enhancing urban biodiversity and its influence on city-dwellers: An experiment. Biol. Conserv. 2014, 171, 82–90. [Google Scholar] [CrossRef]
- Gardner, T.A.; Barlow, J.; Araujo, I.S.; Ávila-Pires, T.C.; Bonaldo, A.B.; Costa, J.E.; Esposito, M.C.; Ferreira, L.V.; Hawes, J.; Hernandez, M.I.M.; et al. The cost-effectiveness of biodiversity surveys in tropical forests. Ecol. Lett. 2008, 11, 139–150. [Google Scholar] [CrossRef]
- Gardner, T.A.; Barlow, J.; Parry, L.W.; Peres, C.A. Predicting the uncertain future of tropical forest species in a data vacuum. Biotropica 2007, 39, 25–30. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Daniel, T.C.; Muhar, A.; Arnberger, A.; Aznar, O.; Boyd, J.W.; Chan, K.M.; Costanza, R.; Elmqvist, T.; Flint, C.G.; Gobster, P.H.; et al. Contributions of cultural services to the ecosystem services agenda. Proc. Natl. Acad. Sci. USA 2012, 109, 8812–8819. [Google Scholar] [CrossRef] [Green Version]
- Silva, C. Community Perceptions of Ecosystem Services and Human Well-Being from Tree Plantations in Argentina. Master’s Thesis, Northern Arizona University, Flagstaff, AZ, USA, 2016. [Google Scholar]
- Córdoba, D.; Selfa, T.; Abrams, J.B.; Sombra, D. Family farming, agribusiness and the state: Building consent around oil palm expansion in post-neoliberal Brazil. J. Rural Stud. 2018, 57, 147–156. [Google Scholar] [CrossRef]
- Pischke, E.C.; Rouleau, M.D.; Halvorsen, K.E. Public perceptions towards oil palm cultivation in Tabasco, Mexico. Biomass Bioenergy 2018, 112, 1–10. [Google Scholar] [CrossRef]
- Pischke, E.C.; Azahara Mesa-Jurado, M.; Eastmond, A.; Abrams, J.; Halvorsen, K.E. Community perceptions of socioecological stressors and risk-reducing strategies in Tabasco, Mexico. J. Environ. Stud. Sci. 2018, 8, 441–451. [Google Scholar] [CrossRef]
- Banerjee, A.; Schelly, C.; Halvorsen, K.E. Understanding public perceptions of wood-based electricity production in Wisconsin, United States: The place-based dynamics of social representations. Environ. Sociol. 2017, 3, 381–393. [Google Scholar] [CrossRef]
- Banerjee, A.; Schelly, C.L.; Halvorsen, K.E. Constructing a sustainable bioeconomy: Multi-scalar perceptions of sustainability. In World Sustainability Series; Springer: Berlin/Heidelberg, Germany, 2018; pp. 355–374. [Google Scholar]
- Barnett, J. Addressing Policy Challenges to Woody Biopower Production: Social Acceptance, Biomass Certification and Limited Policy Support. Ph.D. Thesis, Michigan Technological University, Houghton, MI, USA, 2018. [Google Scholar]
- Abrams, J.; Nielsen, E.; Diaz, D.; Selfa, T.; Adams, E.; Dunn, J.L.; Moseley, C. How do states benefit from nonstate governance? Evidence from forest sustainability certification. Glob. Environ. Polit. 2018, 18, 66–85. [Google Scholar] [CrossRef]
- Dunn, J. Wisconsin Non-Industrial Private Forest Landowners and Wood-Based Bioenergy. PhD Thesis, Michigan Technological University, Houghton, MI, USA, 2018. [Google Scholar]
- Roth, A.M. Retention of Canopy Trees as Biological Legacies for Balancing Woody-Biomass Production and Biodiversity in Managed Aspen Forests of the Great Lakes Region. Ph.D. Thesis, Michigan Technological University, Houghton, MI, USA, 2012. [Google Scholar]
- Roth, A.M.; Flaspohler, D.J.; Webster, C.R. Legacy tree retention in young aspen forest improves nesting habitat quality for Golden-winged Warbler (Vermivora chrysoptera). For. Ecol. Manag. 2014, 321, 61–70. [Google Scholar] [CrossRef]
- De Groot, R.S.; Alkemade, R.; Braat, L.; Hein, L.; Willemen, L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making. Ecol. Complex. 2010, 7, 260–272. [Google Scholar] [CrossRef]
- Guerrero, A.M.; Bennett, N.J.; Wilson, K.A.; Carter, N.; Gill, D.; Mills, M.; Ives, C.D.; Selinske, M.J.; Larrosa, C.; Bekessy, S.; et al. Achieving the promise of integration in social-ecological research: A review and prospectus. Ecol. Soc. 2018, 23, 38. [Google Scholar] [CrossRef]
- Cubbage, F.; Koesbandana, S.; Mac Donagh, P.; Rubilar, R.; Balmelli, G.; Olmos, V.M.; De La Torre, R.; Murara, M.; Hoeflich, V.A.; Kotze, H.; et al. Global timber investments, wood costs, regulation, and risk. Biomass Bioenergy 2010, 34, 1667–1678. [Google Scholar] [CrossRef]
- Cubbage, F.; Mac Donagh, P.; Sawinski, J.; Rubilar, R.; Donoso, P.; Ferreira, A.; Hoeflich, V.; Olmos, V.M.; Ferreira, G.; Balmelli, G.; et al. Timber investment returns for selected plantations and native forests in South America and the southern United States. New For. 2007, 33, 237–255. [Google Scholar] [CrossRef]
- FAO. Global Forest Resources Assessment 2015; UN Food and Agriculture Organization: Rome, Italy, 2015. [Google Scholar]
- Ministerio de Cultura y Comunicación. En Entre Ríos se produce energía alternativa a partir de desechos de la madera—Noticias—Secretaría de Comunicación de la Provincia de Entre Ríos. 2015. Available online: http://www.entrerios.gob.ar/noticias/nota.php?id=41393 (accessed on 5 January 2016).
- Benami, E.; Curran, L.M.; Cochrane, M.; Venturieri, A.; Franco, R.; Kneipp, J.; Swartos, A. Oil palm land conversion in Pará, Brazil, from 2006-2014: Evaluating the 2010 Brazilian Sustainable Palm Oil Production Program. Environ. Res. Lett. 2018, 13, 34037. [Google Scholar] [CrossRef] [Green Version]
- Córdoba, D.; Juen, L.; Selfa, T.; Peredo, A.M.; de Montag, L.F.A.; Sombra, D.; Santos, M.P.D. Understanding local perceptions of the impacts of large-scale oil palm plantations on ecosystem services in the Brazilian Amazon. For. Policy Econ. 2019, 109, 102007. [Google Scholar] [CrossRef]
- Córdoba, D.; Chiappe, M.; Abrams, J.; Selfa, T. Fuelling Social Inclusion? Neo-extractivism, State-Society Relations and Biofuel Policies in Latin America’s Southern Cone. Dev. Chang. 2018, 49, 63–88. [Google Scholar] [CrossRef]
- Pacheco, P.; Gnych, S.; Dermawan, A.; Komarudin, H.; Okarda, B. The Palm Oil Global Value Chain: Implications for Economic Growth and Social and Environmental Sustainability; Center for International Forestry Research (CIFOR): Bogor, Indonesia, 2017. [Google Scholar]
- Monteiro De Carvalho, C.; Silveira, S.; La Rovere, E.L.; Iwama, A.Y. Deforested and degraded land available for the expansion of palm oil for biodiesel in the state of Pará in the Brazilian Amazon. Renew. Sustain. Energy Rev. 2015, 44, 867–876. [Google Scholar] [CrossRef]
- Moreira, J.R.; Goldemberg, J. Oil palm deserves government attention in Brazil. Environ. Res. Lett. 2015, 10, 71002. [Google Scholar] [CrossRef]
- Embrapa. Zoneamento Agroecológico, Produção e Manejo da Cultura de Palma de Óleo na Amazônia (Agro-Ecological Zoning, Production and Management for Growing Oil Palm in the Amazon); Embrapa: Rio de Janeiro, Brazil, 2010. [Google Scholar]
- Nahum, J.S.; Santos, C.B. Impactos socioambientais da dendeicultura em comunidades tradicionais na Amazônia paraense. Rev. ACTA Geográfica 2013, 63–80. [Google Scholar] [CrossRef]
- Abrams, J.; Pischke, E.C.; Mesa-Jurado, M.A.; Eastmond, A.; Silva, C.A.; Moseley, C. Between Environmental Change and Neoliberalism: The Effects of Oil Palm Production on Livelihood Resilience. Soc. Nat. Resour. 2019, 32, 548–565. [Google Scholar] [CrossRef]
- Castellanos-Navarrete, A.; Jansen, K. Oil palm expansion without enclosure: Smallholders and environmental narratives. J. Peasant Stud. 2015, 42, 791–816. [Google Scholar] [CrossRef]
- Isaac-Márquez, R.; Valladares, J.L.S.; Spencer, A.E.; Arcipreste, M.E.A.; Aguilar, M.A.A.; Márquez, A.P.I.; González, M.C.S. Impactos Sociales y Ambientales de la Palma de Aceite: Perspectiva de los Campesinos en Campeche, México. J. Lat. Am. Geogr. 2016, 15, 123–146. [Google Scholar] [CrossRef]
- Castellanos-Navarrete, A.; Jansen, K. Is Oil Palm Expansion a Challenge to Agroecology? Smallholders Practising Industrial Farming in Mexico. J. Agrar. Chang. 2018, 18, 132–155. [Google Scholar] [CrossRef] [Green Version]
- Barbose, G.; Bird, L.; Heeter, J.; Flores-Espino, F.; Wiser, R. Costs and benefits of renewables portfolio standards in the United States. Renew. Sustain. Energy Rev. 2015, 52, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Ebers, A.; Malmsheimer, R.W.; Volk, T.A.; Newman, D.H. Inventory and classification of United States federal and state forest biomass electricity and heat policies. Biomass Bioenergy 2016, 84, 67–75. [Google Scholar] [CrossRef]
- Devine-Wright, P. Reconsidering Public Attitudes and Public Acceptance of Renewable Energy Technologies: A Critical Review; School of Environment and Development, University of Manchester: Manchester, UK, 2007. [Google Scholar]
- Farhar, B.C. Willingness to Pay for Electricity from Renewable Resources: A Review of Utility Market Research. Renew. Energy Lab. 1999, 550, 1–20. [Google Scholar]
- Alban, D.; Perala, D.A.; Jurgensen, M.F.; Ostry, M.E.; Probst, J.R. Aspen Ecosystem Properties in the Upper Great Lakes; North Central Forest Experiment Station, Forest Service—US Department of Agriculture: St. Paul, MN, USA, 1991. [Google Scholar]
- Blyth, J.E.; Smith, W.B. Pulpwood Production in the North Central Region by County, 1987; Resource Bulletin NC-111; U.S. Department of Agriculture, Forest Service, North Central Forest Experiment Station: St. Paul, MN, USA, 1989.
- Cole, K.L.; Davis, M.B.; Stearns, F.; Guntenspergen, G.; Walker, K. Historical landcover changes in the Great Lakes region. In Perspectives on the Land Use History of North. America; Sisk, T.D., Ed.; U.S. Geological Survey, Biological Resources Division: Reston, VA, USA, 1999; pp. 43–50. [Google Scholar]
- White, M.A.; Mladenoff, D.J. Old-growth forest landscape transitions from pre-European settlement to present. Landsc. Ecol. 1994, 9, 191–205. [Google Scholar] [CrossRef]
- Schulte, L.A.; Mladenoff, D.J.; Crow, T.R.; Merrick, L.C.; Cleland, D.T. Homogenization of northern U.S. Great Lakes forests due to land use. Landsc. Ecol. 2007, 22, 1089–1103. [Google Scholar] [CrossRef] [Green Version]
- Niemi, G.J.; Hanowski, J.M. Relationships of Breeding Birds to Habitat Characteristics in Logged Areas. J. Wildl. Manag. 1984, 48, 438–443. [Google Scholar] [CrossRef]
- Jarvi, G.M.; Knowlton, J.L.; Phifer, C.C.; Roth, A.M.; Webster, C.R.; Flaspohler, D.J. Avian Community Response to Short-rotation Aspen Forest Management. Northeast. Nat. 2018, 25, 308–318. [Google Scholar] [CrossRef]
- Niemi, G.J.; Howe, R.W.; Sturtevant, B.R.; Parker, L.R.; Grinde, A.R.; Danz, N.P.; Nelson, M.D.; Zlonis, E.J.; Walton, N.G.; Giese, E.E.G.; et al. Analysis of Long-Term Forest Bird Monitoring Data from National Forests of the Western Great Lakes Region; Gen. Tech. Rep. NRS-159; US Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2016.
- Moskát, C.; Szekely, T. Habitat distribution of breeding birds in relation to forest succession. Folia Zool. 1989, 38, 363–376. [Google Scholar]
- Hobson, K.A.; Bayne, E. The effects of stand age on avian communities in aspen-dominated forests of central Saskatchewan, Canada. For. Ecol. Manag. 2000, 136, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Marsden, S.J.; Whiffin, M.; Galetti, M. Bird diversity and abundance in forest fragments and Eucalyptus plantations around an Atlantic forest reserve, Brazil. Biodivers. Conserv. 2001, 10, 737–751. [Google Scholar] [CrossRef]
- Piratelli, A.; Blake, J.G. Bird communities of the southeastern Cerrado Region, Brazil. Ornitol. Neotrop. 2006, 17, 213–225. [Google Scholar]
- Barlow, J.; Mestre, L.A.M.; Gardner, T.A.; Peres, C.A. The value of primary, secondary and plantation forests for Amazonian birds. Biol. Conserv. 2007, 136, 212–231. [Google Scholar] [CrossRef]
- Calviño-Cancela, M. Effectiveness of eucalypt plantations as a surrogate habitat for birds. For. Ecol. Manag. 2013, 310, 692–699. [Google Scholar] [CrossRef]
- Dias, R.A.; Bastazini, V.A.G.; Gonçalves, M.S.S.; Bonow, F.C.; Müller, S.C. Alterações na composição de comunidades de aves relacionadas ao florestamento de campos temperados no Sudeste da América do su. Iheringia Ser. Zool. 2013, 103, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Simonetti, J.A.; Grez, A.A.; Estades, C.F. Providing Habitat for Native Mammals through Understory Enhancement in Forestry Plantations. Conserv. Biol. 2013, 27, 1117–1121. [Google Scholar] [CrossRef]
- Millan, C.H.; Develey, P.F.; Verdade, L.M. Stand-level management practices increase occupancy by birds in exotic Eucalyptus plantations. For. Ecol. Manag. 2015, 336, 174–182. [Google Scholar] [CrossRef]
- Furumo, P.R.; Aide, T.M. Characterizing commercial oil palm expansion in Latin America: Land use change and trade. Environ. Res. Lett. 2017, 12, 024008. [Google Scholar] [CrossRef]
- Koh, L.P.; Wilcove, D.S. Is oil palm agriculture really destroying tropical biodiversity? Conserv. Lett. 2008, 1, 60–64. [Google Scholar] [CrossRef]
- Almeida, S.M.; Silva, L.C.; Cardoso, M.R.; Cerqueira, P.V.; Juen, L.; Santos, M.P.D. The effects of oil palm plantations on the functional diversity of Amazonian birds. J. Trop. Ecol. 2016, 32, 510–525. [Google Scholar] [CrossRef]
- Lees, A.C.; Moura, N.G.; de Almeida, A.S.; Vieira, I.C.G. Poor Prospects for Avian Biodiversity in Amazonian Oil Palm. PLoS ONE 2015, 10, e0122432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prescott, G.W.; Gilroy, J.J.; Haugaasen, T.; Medina Uribe, C.A.; Foster, W.A.; Edwards, D.P. Reducing the impacts of Neotropical oil palm development on functional diversity. Biol. Conserv. 2016, 197, 139–145. [Google Scholar] [CrossRef]
- Knowlton, J.L.; Phifer, C.C.; Cerqueira, P.V.; Barro, F.D.C.; Oliveira, S.L.; Fiser, C.M.; Becker, N.M.; Cardoso, M.R.; Flaspohler, D.J.; Dantas Santos, M.P. Oil Palm Plantations Affect Movement Behavior of a Key Member of Mixed-Species Flocks of Forest Birds in Amazonia, Brazil. Trop. Conserv. Sci. 2017, 10, 194008291769280. [Google Scholar] [CrossRef] [Green Version]
- Molina, M.; Bohórquez, K. Bird Diversity: A Potential Indicator of Ecologic Sustainability in Agroecosystems at the South of Lake Maracaibo. Boletín Del Cent. Investig. Biológicas 2013, 47, 259–279. [Google Scholar]
- Cajas-Castillo, J.O.; Cóbar-Carranza, A.J.; Carlos, R.; Cruz, Á.-S.; Kraker-Castañeda, C.; Quiñónez-Guzmán, J.M. Diversidad de aves de sotobosque en bosques tropicales, áreas de regeneración natural y cultivos de palma africana en humedales del Lago de Izabal, Guatemala. Ornitol. Neotrop. 2015, 26, 1–12. [Google Scholar]
- Israel, G. Advances in Survey and Data Analysis Methods for Rural Social Scientists: An Introduction. J. Rural Soc. Sci. 2016, 31, 1. [Google Scholar]
- Quattrocchio, M.E.; Borromei, A.M.; Deschamps, C.M.; Grill, S.C.; Zavala, C.A. Landscape evolution and climate changes in the Late Pleistocene-Holocene, southern Pampa (Argentina): Evidence from palynology, mammals and sedimentology. Quat. Int. 2008, 181, 123–138. [Google Scholar] [CrossRef]
- INTA, Instituto Nacional de Tecnologı´a Agropecuaria. Estatsticas agroclimaticas mensuales eea Concordia INTA (Serie historica 1969–2007). 2015. Available online: http://inta.gob.ar/documentos/estadisticas-agroclimaticasde-concordia/at_multi_download/file/INTA_Concordia_Estadisticas_agroclimaticas.pdf (accessed on 8 August 2015).
- Brandão, F.; Schoneveld, G.C. The State of Oil Palm Development in the Brazilian Amazon: Trends, Value Chain Dynamics, and Business Models Governing Oil Palm Landscapes for Sustainability; Africa-China Informal Resource Trade (ACIRT): Bogor, Indonesia, 2015. [Google Scholar]
- Gascon, C.; Bierregaard, R.O., Jr.; Laurance, W.F.; Rankin-de-Merona, J. Deforestation and forest fragmentation in the Amazon. In Lessons from Amazonia: The Ecology and Conservation of a Fragmented Forest; Bierregaard, R.O., Jr., Gascon, C., Lovejoy, T.E., Mesquita, R., Eds.; Yale University Press: New Haven, CT, USA; London, UK, 2001; pp. 22–30. [Google Scholar]
- Haer, T.; Botzen, W.J.W.; Zavala-Hidalgo, J.; Cusell, C.; Ward, P.J. Economic evaluation of climate risk adaptation strategies: Cost-benefit analysis of flood protection in Tabasco, Mexico. Atmosfera 2017, 30, 101–120. [Google Scholar] [CrossRef]
- West, R.; Psuty, N. Las Tierras Bajas de Tabasco en el Sureste de México; No. 972.726 W4.; Gobierno del Estado de Tabasco: Villahermosa, Tabasco, Mexico, 1985.
- Van Der Wal, H.; Pea-Lvarez, B.; Arriaga-Weiss, S.L.; Hernndez-Daums, S. Species, functional groups, and habitat preferences of birds in five agroforestry classes in Tabasco, Mexico. Wilson J. Ornithol. 2012, 124, 558–571. [Google Scholar] [CrossRef]
- Moo Culebro, L.Y.; Knowlton, J.L.; Flaspohler, D.J.; Arriaga-Weiss, S.L.; Mata-Zayas, E.E. Avifauna associated with African palm plantations, secondary vegetation and surrounding matrix of the Sierra Tabasquena. Unpublished.
- NOAA, National Oceanic and Atmospheric Administration. Climate Data Online. National Centers for Environmental Information; NOAA: Ashville, NC, USA, 2016.
- Hutto, R.L.; Pletschet, S.M.; Hendricks, P. A Fixed-radius Point Count Method for Nonbreeding and Breeding Season Use. Auk 1986, 103, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Hillis, D.M.; Mustoe, S. Bird Census Techniques, 2nd ed.; Academic Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Lawrence Erlbaum Associates: Mahwah, NJ, USA, 1988. [Google Scholar]
- Fernández-Llamazares, Á.; Díaz-Reviriego, I.; Guèze, M.; Cabeza, M.; Pyhälä, A.; Reyes-García, V. Local perceptions as a guide for the sustainable management of natural resources: Empirical evidence from a small-scale society in Bolivian Amazonia. Ecol. Soc. 2016, 21, 2. [Google Scholar] [CrossRef] [PubMed]
- Silva-Andrade, H.L.; de Andrade, L.P.; Muniz, L.S.; Telino-Júnior, W.R.; Albuquerque, U.P.; Lyra-Neves, R.M. Do Farmers Using Conventional and Non-Conventional Systems of Agriculture Have Different Perceptions of the Diversity of Wild Birds? Implications for Conservation. PLoS ONE 2016, 11, e0156307. [Google Scholar] [CrossRef]
- King, E.; Cavender-Bares, J.; Balvanera, P.; Mwampamba, T.H.; Polasky, S. Trade-offs in ecosystem services and varying stakeholder preferences: Evaluating conflicts, obstacles, and opportunities. Ecol. Soc. 2015, 20, 25. [Google Scholar] [CrossRef] [Green Version]
- Polasky, S.; Nelson, E.; Camm, J.; Csuti, B.; Fackler, P.; Lonsdorf, E.; Montgomery, C.; White, D.; Arthur, J.; Garber-Yonts, B.; et al. Where to put things? Spatial land management to sustain biodiversity and economic returns. Biol. Conserv. 2008, 141, 1505–1524. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.P.; Gorddard, R.; House, A.P.N.; McIntyre, S.; Prober, S.M. Biodiversity and agriculture: Production frontiers as a framework for exploring trade-offs and evaluating policy. Environ. Sci. Policy 2012, 23, 85–94. [Google Scholar] [CrossRef]
- Ghazoul, J.; Hasanah, N.; Komarudin, H.; Dray, A. Beyond oil palm: Perceptions of local communities of environmental change ETH Library. Front. For. Glob. Chang. 2019, 2, 41. [Google Scholar]
- Azhar, B.; Lindenmayer, D.B.; Wood, J.; Fischer, J.; Manning, A.; McElhinny, C.; Zakaria, M. The conservation value of oil palm plantation estates, smallholdings and logged peat swamp forest for birds. For. Ecol. Manag. 2011, 262, 2306–2315. [Google Scholar] [CrossRef]
- Razak, S.A.; Saadun, N.; Azhar, B.; Lindenmayer, D.B. Smallholdings with high oil palm yield also support high bird species richness and diverse feeding guilds. Environ. Res. Lett. 2020, 15, 094031. [Google Scholar] [CrossRef]
- Kaye-Zwiebel, E.; King, E. Kenyan pastoralist societies in transition: Varying perceptions of the value of ecosystem services. Ecol. Soc. 2014, 19, 17. [Google Scholar] [CrossRef] [Green Version]
- Adams, W.M.; Brockington, D.; Dyson, J.; Vira, B. Managing Tragedies: Understanding Conflict over Common Pool Resources. Science 2003, 302, 1915–1916. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.C.; Pacheco, J.M. Risk of collective failure provides an escape from the tragedy of the commons. Proc. Natl. Acad. Sci. USA 2011, 108, 10421–10425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baars, R.M.T.; Aptidon, S.M. Pastoralists’ perceptions of rangeland degradation in eastern Ethiopia. Nomadic Peoples 2002, 6, 144–157. [Google Scholar] [CrossRef]
- Baird, T.D.; Leslie, P.W.; McCabe, J.T. The effect of wildlife conservation on local perceptions of risk and behavioral response. Hum. Ecol. 2009, 37, 463–474. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Country | Community or State/Province | Feedstock Type | Important/ Very Important | Mean † | Median | Range | SD | N |
|---|---|---|---|---|---|---|---|---|
| Argentina | Entre Ríos | Eucalyptus/Pine | 90% | 4.5 | 5 | 4 | 0.7 | 176 |
| Brazil | Pará | Oil Palm | 86% | 4.5 | 5 | 4 | 0.8 | 212 |
| Mexico | Tabasco | Oil Palm | 99% | 4.6 | 5 | 2 | 0.5 | 130 |
| USA | Wisconsin | Aspen/Northern Hardwood Forest | 83% | 4.3 | 5 | 4 | 0.9 | 288 |
| Country | Community or State/Province | Feedstock Type | Support Expansion | Mean † | Median | Range | SD | N |
|---|---|---|---|---|---|---|---|---|
| Argentina | Entre Ríos | Eucalyptus/Pine | 55% | 3.3 | 4 | 4 | 1.4 | 181 |
| Brazil | Pará | Oil Palm | 60% | 3.5 | 4 | 4 | 1.3 | 208 |
| Mexico | Tabasco | Oil Palm | 60% | 3.5 | 4 | 4 | 1.0 | 128 |
| USA | Wisconsin | Aspen/ Northern Hardwood Forest | 48% | 3.3 | 4 | 4 | 1.1 | 278 |
| Country | Community or State/Province | Feedstock Type | Yes | Mean † | Median | Range | SD | N |
|---|---|---|---|---|---|---|---|---|
| Argentina | Entre Ríos | Eucalyptus/Pine | 71% | 0.9 | 1 | 1 | 0.7 | 150 |
| Brazil | Pará | Oil Palm | 34% | 0.4 | 0 | 1 | 0.5 | 182 |
| Mexico | Tabasco | Oil Palm | 74% | 0.8 | 1 | 1 | 0.4 | 117 |
| Country | Community or State/Province | Feedstock Type | Negatively/ Very Negatively | Mean † | Median | Range | SD | N |
|---|---|---|---|---|---|---|---|---|
| Argentina | Entre Ríos | Eucalyptus/Pine | 17% | 3.6 | 4 | 4 | 1.2 | 173 |
| Brazil | Pará | Oil Palm | 52% | 2.7 | 2 | 4 | 1.4 | 202 |
| Mexico | Tabasco | Oil Palm | 19% | 3.7 | 4 | 4 | 1.1 | 125 |
| USA | Wisconsin | Aspen/Northern Hardwood Forest | 45% | 2.8 | 3 | 4 | 1.3 | 282 |
| Country | Ranking of Perception of Negative Feedstock Impact on Birds | Perception of Feedstock Expansion Impacts Mean (Table 4) | Respondents Saying Feedstock Expansion Negatively Impacts Birds | Feedstock Versus Native Forest Bird Habitat Ecological Effect Size Difference | Ranking of Ecological Effect Size Difference |
|---|---|---|---|---|---|
| Argentina | 4 | 3.6 | 17% | 6.209 | 2 |
| Brazil | 1 | 2.7 | 52% | 9.619 | 1 |
| Mexico | 3 | 3.7 | 19% | 2.434 | 3 |
| USA | 2 | 2.8 | 45% | 1.616 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knowlton, J.L.; Halvorsen, K.E.; Flaspohler, D.J.; Webster, C.R.; Abrams, J.; Almeida, S.M.; Arriaga-Weiss, S.L.; Barnett, B.; Cardoso, M.R.; Cerqueira, P.V.; et al. Birds and Bioenergy within the Americas: A Cross-National, Social–Ecological Study of Ecosystem Service Tradeoffs. Land 2021, 10, 258. https://doi.org/10.3390/land10030258
Knowlton JL, Halvorsen KE, Flaspohler DJ, Webster CR, Abrams J, Almeida SM, Arriaga-Weiss SL, Barnett B, Cardoso MR, Cerqueira PV, et al. Birds and Bioenergy within the Americas: A Cross-National, Social–Ecological Study of Ecosystem Service Tradeoffs. Land. 2021; 10(3):258. https://doi.org/10.3390/land10030258
Chicago/Turabian StyleKnowlton, Jessie L., Kathleen E. Halvorsen, David J. Flaspohler, Christopher R. Webster, Jesse Abrams, Sara M. Almeida, Stefan L. Arriaga-Weiss, Brad Barnett, Maíra R. Cardoso, Pablo V. Cerqueira, and et al. 2021. "Birds and Bioenergy within the Americas: A Cross-National, Social–Ecological Study of Ecosystem Service Tradeoffs" Land 10, no. 3: 258. https://doi.org/10.3390/land10030258